噬菌体展示技术是一种强大的体外筛选平台,能够将多肽或蛋白质(如抗体)与其编码的遗传物质(DNA)物理连接在一起,从而实现从庞大文库中高效淘选、富集出能与特定靶标结合的分子。它在纳米抗体开发中扮演着核心角色,极大地加速了筛选过程。
一、 技术核心原理
基因型与表型关联:将编码目标蛋白(如纳米抗体VHH片段)的外源基因,插入噬菌体某个外壳蛋白的基因序列中。
融合展示:在感染宿主细胞后,噬菌体利用自身的复制和组装系统,将外源基因表达为融合蛋白,并使其呈现在新生噬菌体颗粒的表面。被展示的蛋白通常能保持其天然构象和生物活性。
生物淘选:

结合:将展示有随机蛋白库的噬菌体群体与固定的靶抗原(如包被在酶标板上的蛋白质)共同孵育。
洗脱:洗去未结合或弱结合的噬菌体。
洗脱与扩增:将特异性结合的噬菌体洗脱下来,并感染大肠杆菌进行扩增,得到富集了潜在结合子的子代噬菌体库。
循环:重复上述“吸附-洗脱-扩增”过程(通常3-4轮),可指数级富集出高亲和力、高特异性的克隆。
二、 主要噬菌体展示系统及其特点
根据所用噬菌体载体的不同,系统各有优劣,适用于不同场景。
系统类型 | 代表载体 | 基因组特点 | 展示特点与优势 | 主要适用场景 |
丝状噬菌体 | M13, fd, f1 | 单链环状DNA | 最常用系统。通过pIII蛋白(低拷贝,3-5个,展示大蛋白)或pVIII蛋白(高拷贝,~2700个,展示短肽)融合。不裂解宿主,可连续生产,滴度高。 | 抗体、蛋白、多肽文库的构建与筛选。 |
T7噬菌体 | T7 | 线性双链DNA | 烈性噬菌体,组装在胞内,通过裂解释放。复制速度极快(整个周期2-3小时),筛选周期短。可展示非分泌型或对细菌有毒的蛋白。 | 需快速筛选的场景,展示胞内蛋白或毒性蛋白。 |
T4噬菌体 | T4 | 大型线性双链DNA | 拥有Soc和Hoc两个非必需衣壳蛋白位点,可同时或分别展示两种外源蛋白,展示拷贝数高(可达数百个)。组装在胞内,可展示大分子复合物。 | 展示大型蛋白复合物、多价展示、疫苗研发。 |
λ噬菌体 | λ | 双链DNA | 在宿主细胞内组装,具有特征性的二十面体头部。主要融合位点为尾部蛋白(如pV)。适于展示有活性的大分子或细胞毒性蛋白。 | 展示酶、细胞表面受体配体等。 |
三、 M13展示系统与“噬菌粒-辅助噬菌体”系统
由于M13最为常用,其技术体系也发展得最为成熟和完善。
1. 经典的M13噬菌体载体:
外源基因直接插入改造后的M13基因组(如M13mp系列)。虽然直接,但载体较大,克隆效率相对较低,且外源基因过长可能影响噬菌体稳定性。

2. “噬菌粒-辅助噬菌体”系统:
为了克服经典载体的缺点,此系统成为目前的主流。
一个融合基因:外源蛋白基因 + 噬菌体外壳蛋白(gIII 或 gVIII)基因片段。
质粒复制起点。
抗生素抗性基因(如Amp⁺)。
M13噬菌体的复制起点。
1. 将含有外源基因的噬菌粒转化进大肠杆菌。
2. 用辅助噬菌体超感染该细菌。
3. 辅助噬菌体提供所有的病毒蛋白,但由于其自身DNA复制效率低,这些蛋白会“优先”识别并包装复制效率更高的噬菌粒的单链DNA。
4. 最终,子代噬菌体颗粒携带的是噬菌粒DNA,并在表面展示由噬菌粒编码的融合蛋白。辅助噬菌体自身的基因组很少被包装。
此系统的优势:
ü 高稳定性:噬菌粒作为质粒,在细菌中更稳定,可携带更大、更复杂的外源片段。
ü 高克隆效率:小质粒更易于进行分子克隆操作。
ü 灵活性:同一噬菌粒可用不同辅助噬菌体拯救,且可方便地切换展示格式(如从噬菌体展示转为可溶性表达)。
总结
噬菌体展示技术通过巧妙的“基因型-表型”关联,建立了高效的体外筛选管道。其中,基于M13的“噬菌粒-辅助噬菌体”系统因其稳定、高效、灵活的特性,已成为抗体(尤其是纳米抗体)筛选的金标准平台。研究者可根据目标蛋白的特性(大小、毒性、是否需要分泌等)和筛选需求(速度、拷贝数等)选择最合适的展示系统。
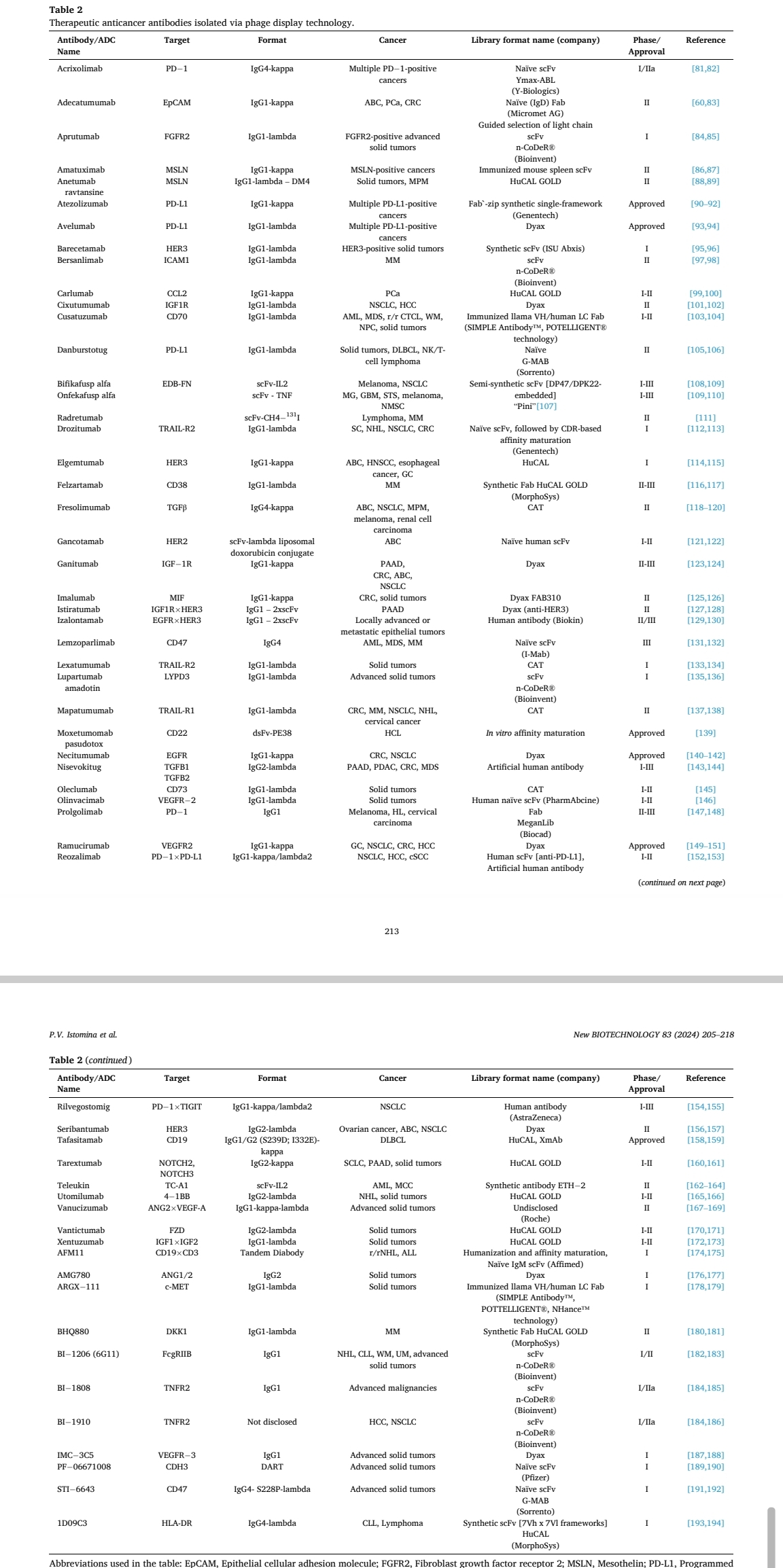
附录:利用噬菌体技术筛选到的药物抗体清单(截止2024年)
参考文献
Istomina PV, Gorchakov AA, Paoin C, Yamabhai M. Phage display for discovery of anticancer antibodies. N Biotechnol. 2024 Nov 25;83:205-218. doi: 10.1016/j.nbt.2024.08.506. Epub 2024 Aug 24. PMID: 39186973.







